生物信息学学习笔记(三)

- CpG位点(CpG sites,CG位点):指DNA的某个区域,其上的碱基序列以胞嘧啶接着鸟嘌呤出现。CpG是“—C—磷酸—G—”的缩写,指磷酸二脂键连接了胞嘧啶和鸟嘌呤,其中C位于5’端而G位于3’端。在CpG位点中的胞嘧啶可以被甲基化为5-甲基胞嘧啶,基因内CpG位点的甲基化会改变基因的表达
顺式调控元件
-
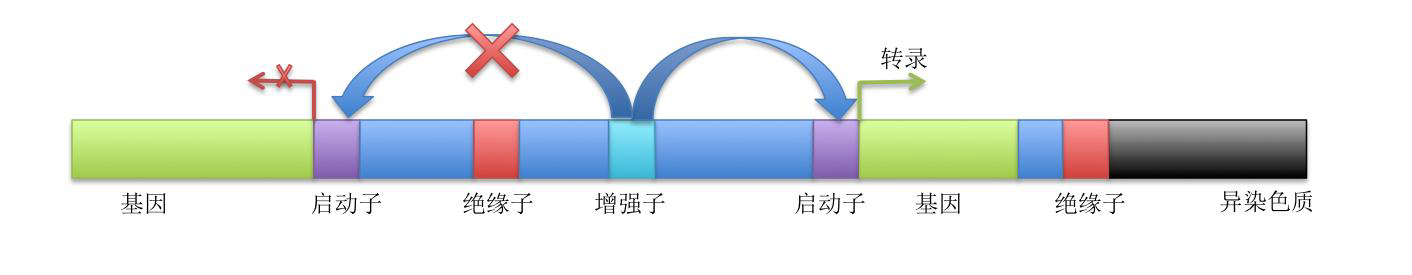
启动子(promoter):指导RNA聚合酶并起始DNA转录的一段DNA序列,该序列往往位于编码基因的上游,可以调控目标基因表达的程度、时间和组织等。通常核心启动子都具有一些特征序列,这些特征序列称为基序(motif),常见的基序有TATA盒(TATA box)等。
-
增强子和沉默子:指DNA分子上可以与特定蛋白相结合从而正向或负向调控基因表达的顺式作用元件。其中增强子与蛋白结合之后,基因的转录水平会增强,其位置可以位于启动子上游、基因内、基因下游或基因区间,甚至于被调控基因不在同一条染色体上,这是由于增强子主要依靠空间作用来调控基因表达。
-
绝缘子:绝缘子是一种长程调控元件,主要有两类。第一类主要功能是阻遏邻近的调控元件如增强子。绝缘子发挥功能时具有方向性,其可以阻断增强子与启动子的作用,从而导致增强子的目标基因表达下调。第二类是异染色质的边界原件。由于异染色质包装折叠较常染色体显得更加紧密,异染色质中的基因很少表达,而第二类绝缘子可以在异染色质的边界阻断异染色质结构的延伸,保证边界另一侧的基因表达不受影响。
-
转录水平的调控是真核基因表达调控的关键环节,染色质对DNase Ⅰ的敏感性与基因转录有关。基因活化使其染色质一呈开放的疏松型构象,更易被DNase Ⅰ降解,形成DNase Ⅰ超敏感位点(DNase Ⅰ hypersensitive site,DHSs).DHSs不仅与染色质的开放度相关,还与启动子、增强子、绝缘子以及沉默子等元件相偶联,并且染色质上具有调控功能的DNA序列、调控蛋白结合位点也与DHSs重叠
-
DHSs是一段跨度为数百碱基对,甲基化程度较低,对DNase Ⅰ具有高度敏感性的染色质区域,可富集转录因子和重要酶类,具有转录调控功能
-
染色质由一系列盘绕着DNA的核小体组成,转录因子在与基因结合时,取代了核小体的位置,baoluchuDNA,使其对DNase Ⅰ的切割更为敏感,在此基础上,利用染色质对DNase Ⅰ的敏感性鉴定活化的调控序列
-
DNase-seq方案包括用DNase Ⅰ优先切割核小体被取代的DNA序列,对生成的短片段进行测序并将其定位到基因组上以鉴定对敏感酶的“染色质开放”区域。一些转录因子的结合位点显示出高度特异性的DNase Ⅰ剪切模式。这种“Dnase 足迹”常被用来检测特定转录因子的结合
-
ATAC-seq(assay for transposase-accesiible chromatin using sequencing):通过Tn5转座酶优先标记和测序核小体之间的DNA。类似于DNase-seq,但是步骤更为简单,所需细胞也更少
-
对DNase-seq片段进行双端测序,发现短片段主要源自于核小体之间的区域,而长片段跨越了核小体。研究指出短片段对鉴定转录因子的结合更有用,长片段可以提供核小体的间隔信息
-
研究人员发现由于DNase Ⅰ和Tn5的切割偏好性,之前许多Dnase印记分析反映的更多是切割偏好而不是转录因子的真实结合情况。
3D基因组
- 不同染色质在细胞核内的分布不是杂乱无章的,而是被限定在特定的“疆域”内,基因相对密集的染色质通常分布在远离核膜的位置,而基因稀疏的染色质则分布在靠近核膜的位置。在基因组上分离的染色质区段在空间上可能被拉到一起共同被转录,形成“转录工厂”
- 基因组区室是指存在于染色质上的相互间隔的连续区域,通常这些区域大小为1~10Mb,每个区域内的点具有相似的全基因组交互模式,根据交互模式的差异,这些区域可以分为两类:A类区室和B类区室。相同类型的区室具有相似的全基因组交互模式,其交互频率要显著高于不同类型区室之间的交互。A类区室往往代表基因富集的常染色质区域,B类区室则对应着基因稀疏的异染色质区域
- 拓扑相关结构域(TAD)则是位于高等动物染色质的更精细的结构单元,TAD被定义为这样一块块连续的染色质区域,区域内部互作强度要显著强于区域之间的互作。每个TAD通常只有200Kb~1Mb。
- 染色质位点的相互作用存在某种固有的模式,而这些模式与特定生物学过程/功能是无关的